Acremonium
The genus Acremonium contains many species; most are saprophytic being isolated from dead plant material and soil.
Several species have been reported as opportunistic pathogens of man and animals, causing mycetoma, mycotic keratitis and onychomycosis, however molecular studies have shown that only Sarocladium kiliense and Acremonium egyptiacum have affected a significant number of patients (Perez-Cantero and Guarro, 2020).
Recently, several Acremonium-like species previously reported as opportunistic pathogens have been transferred to other genera; Xenoacremonium recifei (formerly A. recifei), Neocosmospora falciforme (formerly A. falciforme), Gliomastic roseogrisea (formerly A. roseogriseum), Sarocladium kiliense (formerly A. kiliense) and Sarocladium strictum (formerly A. strictum) (Glenn et al., 1996; Summerbell et al., 2011; Lombard et al., 2015).Several species including A. recifei and A. alabamense are recognised as opportunistic pathogens of man and animals, causing mycetoma, mycotic keratitis and onychomycosis.
RG-2 for species isolated from humans.
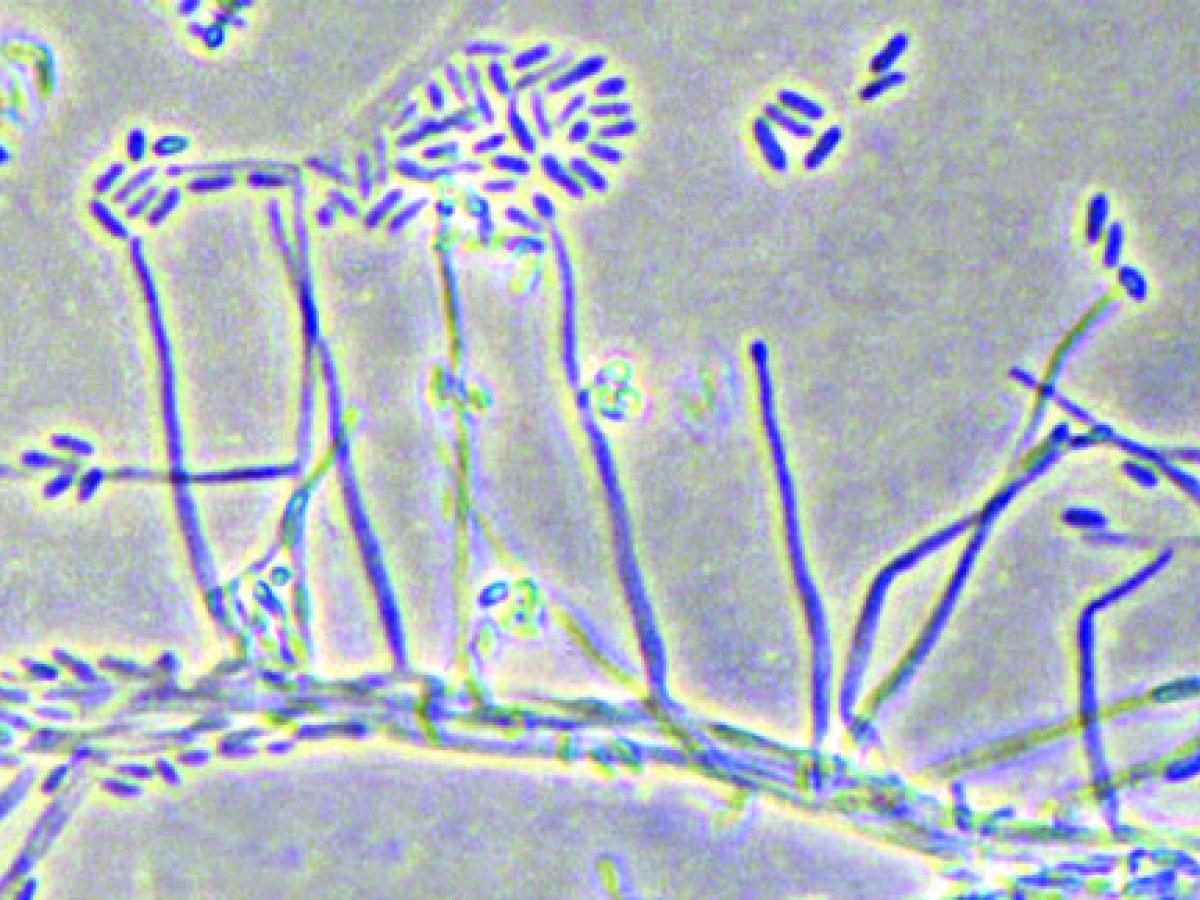
Acremonium sp. showing long awl-shaped phialides producing cylindrical, one-celled conidia mostly aggregated in slimy heads at the apex of each phialide.
Morphological description:
Colonies are usually slow growing, often compact and moist at first, becoming powdery, suede-like or floccose with age, and may be white, grey, pink, rose or orange in colour. Hyphae are fine and hyaline and produce mostly simple awl-shaped erect phialides with inconspicuous collarettes. Conidia are usually one-celled, hyaline or rarely pigmented, globose to cylindrical, and mostly aggregated in slimy heads at the apex of each phialide. Chlamydospores may be present.
Comments:
Microconidial Fusarium isolates may be confused with Acremonium, but they usually grow faster and have colonies with a characteristic fluffy appearance. Phialemonium species differ by having short, tapering phialides, mostly lacking a basal septum. Coniochaeta is characterised by having sessile phialidic collarettes that are formed directly on the hyphae.
Key features:
Hyphomycete with solitary, erect, hyaline, awl-shaped phialides producing single-celled, globose to cylindrical conidia, mostly in slimy heads.
Molecular identification:
Summerbell et al. (2011) revised the genus on the basis of 18S and D1/D2 sequence phylogeny. Sequence based identification may be performed using the D1/D2 or the ITS region. Caution must be exercised though in the interpretation of database sequence comparisons due to the scarcity of database sequences from well-characterised strains, and some sequences may have been attributed to species that have been reclassified (Perdomo et al. 2011a).
| Antifungal | No | 0.016 | 0.03 | 0.06 | 0.125 | 0.25 | 0.5 | 1 | 2 | 4 | ≥8 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| AmB | 10 | 1 | 2 | 1 | 1 | 2 | |||||
| ISAV | 2 | 1 | 1 | ||||||||
| VORI | 10 | 1 | 3 | 4 | 1 | 1 | |||||
| POSA | 9 | 1 | 2 | 1 | 4 | 1 | |||||
| ITRA | 10 | 2 | 1 | 4 | 3 |
References: Gams (1971), Domsch et al. (2007), Samson et al. (1995), de Hoog et al. (2000, 2015), Glenn et al. (1996), Perdomo et al. (2011a), Summerbell et al. (2011).